![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

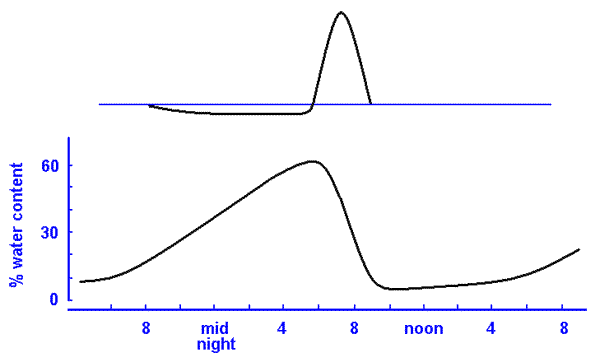
Case StudiesHydration & photosynthesisLichens rely directly on their surroundings for water and tend to match the water levels of their immediate environment. They dry out (and become dormant) as their surroundings dry out and become hydrated when there is sufficient water in the surrounds. Bryophytes are another group of organisms with similar water behaviour and both are examples of poikilohydric organisms. By contrast vascular plants generally have mechanisms which aim to maintain internal water even in dry conditions and are called homoihydric (or homoiohydric). Poikilohydry is very rare amongst the vascular plants and one example is the South American succulent Blossfeldia liliputana, which can lose up to 80% of its internal water. A dry lichen thallus is physiologically inactive. A hydrated lichen thallus will respire, which is to say it will consume stored carbohydrates and produce carbon dioxide in the process. A hydrated lichen thallus that is exposed to the light will respire but also photosynthesize and produce carbohydrates. When a dry thallus is moistened there will be an initial period when respiration outruns photosynthesis. This is obviously true if the moistened thallus is kept in the dark since no photosynthesis can then occur, but it is also true for a well-lit thallus. If a moistened thallus is taken from the dark into the light it will begin to photosynthesize and if photosynthesis is able to continue long enough there will come a time when it will have 'caught up' and replenished the store of carbohydrates that were used during the initial period of respiration. If photosynthesis is able to continue beyond the 'catch up' point additional carbohydrates will be produced and stored within the thallus. In the case of a thallus that was moistened in the light it is clear that the 'catch up' period would be shorter. In many areas lichens often become hydrated in the evening or early morning from water vapour or dew. This means there can be many hours of respiration before the early morning light allows the start of photosynthesis. The rate at which carbon dioxide is being produced or taken up can be measured and the results plotted as a function of time. The graph (below left) shows what such a plot might look like. In this example I will assume there is no additional water input after the fog/dew rehydration. The vertical axis measures the rate of carbon dioxide exchange between thallus and atmosphere. There is a zero point on that axis. A negative number means the rate of production (via respiration) is greater than the rate of uptake (via photosynthesis). A positive number means the rate of uptake is greater than the rate of production. The horizontal axis represents time, with later time to the right. At first the thallus is dry and inactive but at the time indicated by the arrow to the left the thallus has absorbed some water and is beginning to respire. After some time the thallus reaches a steady respiration rate. At the time indicated by the other arrow photosynthesis begins so the black plot line begins to go up. For a while respiration continues to outpace photosynthesis but eventually the black line reaches the zero line. At that point the rate of carbon dioxide production is equal to the rate of carbon dioxide uptake. While the rates are equal there is still net carbon dioxide production so that more stored carbohydrate has been used than has been replaced by photosynthesis. In such plots the vertical axis measures the amount of carbon dioxide per gram (dry weight) of thallus per some unit of time or (especially for crustose species) amount per unit area per unit time. Given that time is measured along the horizontal axis basic calculus shows that total amounts of carbon dioxide production (or uptake) can be obtained by measuring the relevant areas between the plot line and the zero line. In particular the total production of carbon dioxide (per gram or per unit area) between the beginning of hydration and the moment photosynthesis matches respiration is measured by the red area of the right hand figure of the above pair. This amount must be made up during photosynthesis if the reserves used during the earlier respiration are to be replaced. In the plot so far the carbon dioxide exchange level has climbed into the positive range and the total amount of carbon dioxide taken up so far by photosynthesis is shown by the small grey area in the right-hand figure. The grey area is clearly smaller than the red area so there is still some way to go before the carbohydrate reserves used up during respiration have been replaced. You should not think that the production of carbon dioxide during the "red" period has been wasteful or represents some sort of loss to the lichen. The stored carbohydrates will have been used for repair or growth and carbon dioxide is a by-product of that usage. It is no different to a human body where stored reserves are used for maintenance. A lichen thallus (just like a human) can use reserves for a while but if this continues for too long without replenishment the thallus (or human) would die. Hence, too many occurrences of the first scenario of the previous paragraph would be fatal. The above plots show some artificial scenarios in order to explain some basic principles. The next diagram, with four realistic plots, shows the behaviour of Lecanora muralis thalli on four different days when there were different patterns of hydration. The measurements were made at the Würzburg Botanic Garden in Germany. Since I wish to illustrate only the variation in patterns I have not put values along the vertical axis. The scale is linear so you could work out relative rates of photosynthesis should you wish to do so. (B) A thallus was thoroughly wetted by overnight rain but the following day was clear. Carbon dioxide diffuses very slowly through water so a saturated thallus impedes gas exchange and hence also photosynthesis. On this day the saturated thallus did see a slow rise in photosynthesis in the early post-sunrise hours. As the sun rose further and the day warmed up the thallus began to dry out. By late morning the thallus had dried sufficiently to allow much better gas exchange and this explains the sudden spike in the red graph. During the early afternoon the thallus dries out. (C) Overnight rain again produced a saturated thallus so there was only moderate photosynthesis in the early post-sunrise hours. Then, as in case B, there was an increase in photosynthesis once the thallus had dried sufficiently and a sharp rise in the grey graph. Once the thallus started drying the rate of photosynthesis dropped but, before the thallus had dried completely there was light rain. The rain was not enough to saturate the thallus but enough to lead to a rise in photosynthesis. This happened two more times during this day and so you see four peaks in this graph. Of the four days featured in the diagram, this day saw the greatest amount of carbohydrates produced. (D) Overnight rain produced a saturated thallus and rain during the day kept the thallus supersaturated for much of the day and therefore unable to take advantage of the periods of good light. Of the four days shown here this day saw the least amount of carbohydrates produced. The photosynthetic rates of wet thalli vary, with some species able to maintain higher rates of photosynthesis than others. A number of hydrophobic compounds produced by lichens prevent over-saturation and so allow gas exchange (and hence photosynthesis) to continue even in wet environments. In hot, dry, inland environments where rainfall is rare thallus hydration is typically brought about by evening or overnight dew and thallus water content rises through the pre-dawn hours. After sunrise there is a period of no more than about 3 hours during which photosynthesis occurs but typically by late morning the temperature has risen enough to drive lichen thalli back into dormancy. The following pair of graphs shows such a pattern for the fruticose species Ramalina maciformis in Israel's Negev desert during September 26-27 in 1967. The upper, similar to the graphs given above, shows the pattern of carbon dioxide exchange and the lower graph shows thallus water content (as a percentage of dry thallus weight). The horizontal axis applies to both graphs. Over 24 hours the uptake of carbon dioxide was 1.32 milligrams per gram of dry thallus weight and the production via respiration was 0.78, yielding a surplus of 0.54 milligrams of carbon dioxide per gram of dry thallus weight. This means that carbon dioxide production was about 60% of total uptake during photosynthesis. Obviously there will be variation in respiration and photosynthesis depending on external conditions during any particular night-day period but various studies of lichens in a variety of habitats have shown that the 60% found here is neither unusually high nor unusually low. Lichens are not extraordinary in this respect since vascular plants also show significant percentages. That surplus of 0.54 milligrams of carbon dioxide equates to a gain of 0.146 milligrams of carbon per gram of dry thallus weight. If this were a reasonable long term average some arithmetic shows that it would take a thallus about 19 years to double its carbon content. For photosynthesis to occur a thallus needs to be moist. However, you've seen that if a thallus is too moist the rate of photosynthesis will be low and therefore it is clear that there is a moisture content which would promote an optimum rate of photosynthesis. Similarly, though this has not been shown above, there are temperatures and light levels at which photosynthesis would be optimum. The optimum conditions for photosynthesis vary between species. References
|
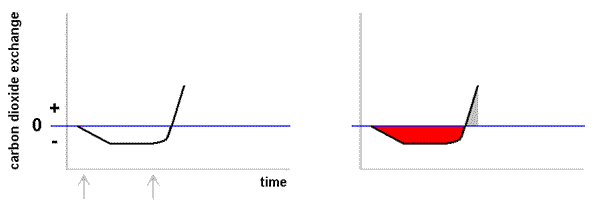
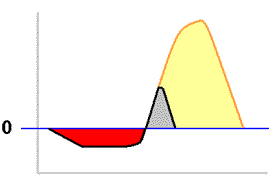 The diagram on the right of this paragraph shows a couple of possible continuations of the plot into the later hours of the day. Suppose that soon after photosynthesis had matched respiration it becomes hot enough for the atmospheric humidity to begin to drop rapidly. Photosynthesis could continue to increase while there was still sufficient water within the thallus. However, being poikilohydric, the thallus would soon begin to dry, the rate of photosynthesis would slow and then decrease as the thallus became drier and drier. Eventually the thallus would become dry enough to become dormant once more. This scenario is shown by the black plot line and the total amount of carbon dioxide that has been taken up during photosynthesis is shown by the grey area. It is clear that this is less than the red area so, in this scenario, the thallus has not replaced the carbohydrate reserves used up by respiration. Suppose instead that the drying out does not start so early and that we see a plot like the one shown by the orange-brown line. In this scenario the total amount of carbon dioxide taken up during photosynthesis is shown by the combined grey and yellow areas, clearly greater than the red area. In the second scenario the reserves have been replaced and additional carbohydrates have been produced, a good outcome for the thallus.
The diagram on the right of this paragraph shows a couple of possible continuations of the plot into the later hours of the day. Suppose that soon after photosynthesis had matched respiration it becomes hot enough for the atmospheric humidity to begin to drop rapidly. Photosynthesis could continue to increase while there was still sufficient water within the thallus. However, being poikilohydric, the thallus would soon begin to dry, the rate of photosynthesis would slow and then decrease as the thallus became drier and drier. Eventually the thallus would become dry enough to become dormant once more. This scenario is shown by the black plot line and the total amount of carbon dioxide that has been taken up during photosynthesis is shown by the grey area. It is clear that this is less than the red area so, in this scenario, the thallus has not replaced the carbohydrate reserves used up by respiration. Suppose instead that the drying out does not start so early and that we see a plot like the one shown by the orange-brown line. In this scenario the total amount of carbon dioxide taken up during photosynthesis is shown by the combined grey and yellow areas, clearly greater than the red area. In the second scenario the reserves have been replaced and additional carbohydrates have been produced, a good outcome for the thallus.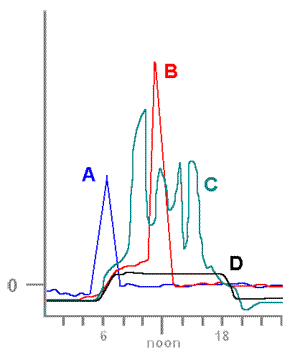 (A) This day was similar to that illustrated by the artificial plots given earlier. A thallus was hydrated by overnight dew, respired in the pre-dawn hours and after sunrise photosynthesis rose to a peak before the lichen started to dry. At that point photosynthesis dropped until the thallus was once more dry.
(A) This day was similar to that illustrated by the artificial plots given earlier. A thallus was hydrated by overnight dew, respired in the pre-dawn hours and after sunrise photosynthesis rose to a peak before the lichen started to dry. At that point photosynthesis dropped until the thallus was once more dry.