![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)


Life cycle
This page will start with the bryophyte...
... life cycle in a nutshell.
A bryophyte spore is haploid. A haploid cell has one set of chromosomes, a diploid cell has two. In humans, the egg and sperm cells are haploid. When they unite, the resulting cell is diploid and the ensuing embryo continues its development as a diploid individual. Each parent has contributed one set of chromosomes.
A bryophyte spore germinates and produces an often algal-like mat, called a protonema (plural protonemata) and the leafy or thalloid stage of the gametophyte develops from the protonemal stage. The protonemata are almost always ephemeral but there are exceptions. Amongst the mosses there is a small number of species in which the protonemata are persistent and the leafy plants are ephemeral. The spore was haploid and so are the resulting protonema and the ensuing leafy or thalloid stages. The male and female sex organs, the antheridia and the archegonia respectively, are produced on the gametophytic plants. Haploid sperm are released from the antheridia and when a haploid sperm reaches a haploid egg in an archegonium the egg is fertilized to produce a diploid cell. This cell will develop into the diploid sporophyte. In a great many bryophytes the tissue in the embryonic sporophyte differentiates. Typically there will be a foot, that anchors the embryonic sporophyte to the supporting gametophyte, and at the end opposite the foot is the part that will develop into the spore capsule. In between there is typically the part that will become the stalk (or seta) that supports the spore capsule, though in a number of bryophytes the seta is short or even non-existent. By the time the sporophyte has matured the spore capsule will contain haploid spores.
The following diagram gives one example of a bryophyte life cycle, that of the moss Funaria hygrometrica.
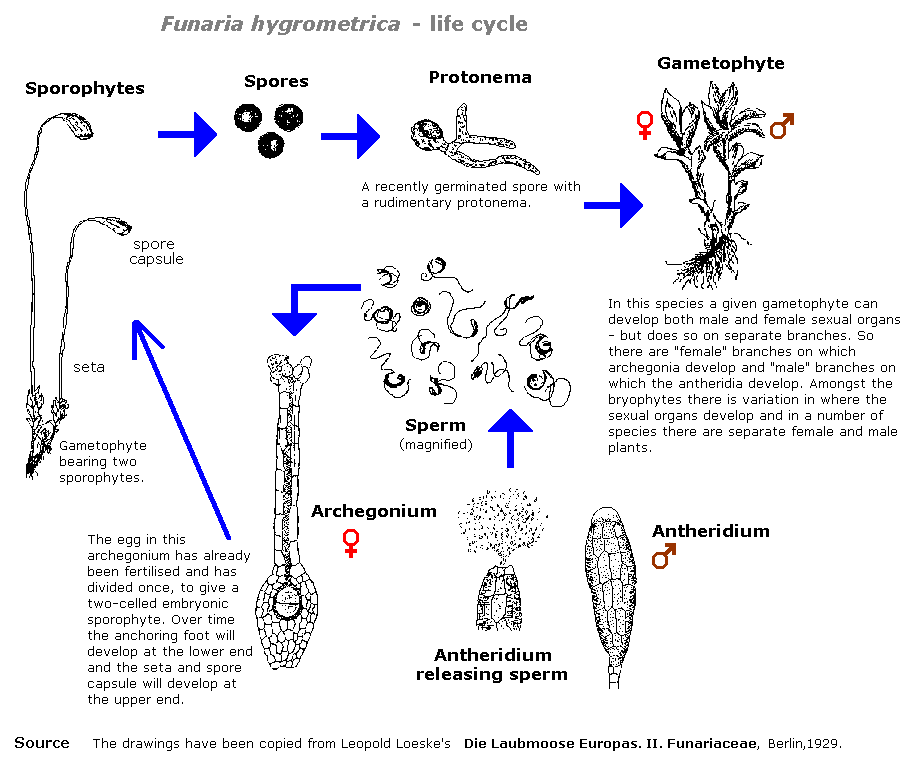
Print this diagram as an A4 PDF
The spores produced in a spore capsule are the result of SEXUAL REPRODUCTION. Bryophytes are adept at VEGETATIVE REPRODUCTION and vegetative propagules often produce protonemata rather than developing directly into the leafy or thalloid gametophytic stage.
The bryophytes are often said to show an alternation of generations. You may see references to the "gametophyte generation" and the "sporophyte generation" alternating. The first thing to note is that "generation" is being used in a way quite different to its every day use. Parents (diploid organisms) refer to their children (also diploid organisms) as the "next generation". The second point worth noting is that many bryophyte gametophytes readily produce offspring without need of sporophytes. The third point to not is that a great many organisms show haploid and diploid states. In humans the haploid state is confined to egg and sperm cells and these are incapable of an independent existence. In the flowering plants the eggs and pollen constitute the haploid state and are also not capable of independent existence. Humans and flowering plants are examples of diploid-dominant organisms. By contrast, in bryophytes the haploid state is capable of independent existence and is typically much longer-lived than the sporophyte state. Bryophytes are examples of haploid-dominant organisms.
The rest of this page is devoted mostly to the subject of gametophyte development, from spore through to the leafy or thallose stage. This page finishes with some interesting web links. There is a separate SPOROPHYTE DEVELOPMENT page.
Spore to protonema
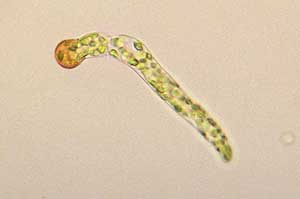 Germinating Funaria spore |
Upon germination a spore typically produces a short, filamentous outgrowth called a germ tube. The photo shows a germinating spore of a moss in the genus Funaria. The spore is the brownish spherical object and a germ tube has grown out from it. The tube shown in this photo is already several cells long and you can also see many green, chlorophyll-filled chloroplasts, in which photosynthesis is carried out. The initial tube lengthens and branches and this photo ![]() shows the filamentous complexes that have developed from three Funaria spores. The early growth in many moss protonema consists of filamentous tubes with colourless walls and with the cross-walls between neighbouring cells being at right-angles to direction of the filament's long axis. This stage is called the chloronema. Later a caulonema typically develops. The growth is still filamentous but now the walls are pigmented (typically reddish-brown) and the cross-walls are obliquely oriented.
shows the filamentous complexes that have developed from three Funaria spores. The early growth in many moss protonema consists of filamentous tubes with colourless walls and with the cross-walls between neighbouring cells being at right-angles to direction of the filament's long axis. This stage is called the chloronema. Later a caulonema typically develops. The growth is still filamentous but now the walls are pigmented (typically reddish-brown) and the cross-walls are obliquely oriented.
The following stylized diagrams show some of the variations.
In the first pair of diagrams the grey moss spore has germinated and produced a chloronema (shown as ![]() , with the black lines indicating the cross-walls between neighbouring cells). Soon after germination you can see branches in the chloronema and in the diagram on the left you can see the development of a caulonema, with its obliquely oriented cross-walls, shown by
, with the black lines indicating the cross-walls between neighbouring cells). Soon after germination you can see branches in the chloronema and in the diagram on the left you can see the development of a caulonema, with its obliquely oriented cross-walls, shown by ![]() . You can also see the beginning of leafy growth (
. You can also see the beginning of leafy growth ( ![]() ) from the caulonema. The chloronema is rich in chloroplasts but the caulonema has relatively few. The leafy stage of the gametophyte typically develops from a caulonema but, as shown by the diagram on the right, in some mosses leafy growth develops from the chloronema. It is possible for a caulonema to give rise to fresh chloronema, as shown by a partially dotted-line chloronema in the diagram on the left.
) from the caulonema. The chloronema is rich in chloroplasts but the caulonema has relatively few. The leafy stage of the gametophyte typically develops from a caulonema but, as shown by the diagram on the right, in some mosses leafy growth develops from the chloronema. It is possible for a caulonema to give rise to fresh chloronema, as shown by a partially dotted-line chloronema in the diagram on the left.

It is important that you understand that all the protonema diagrams on this page simply show the various early stages in gametophyte growth and their relationships to each other. Don't take them as realistic depictions of actual gametophyte development. For example, taken literally, the two diagrams immediately above show the leafy buds developing on a small protonema, seemingly very soon after spore germination. In fact a moss protonema can be very extensive before the leafy buds begin developing.
 In most mosses the protonema is filamentous but in a few genera it is thalloid. The diagram (right) shows an example from the genus Sphagnum. There is a short, filamentous chloronema but it soon develops into a thallose form. The thallose area, though composed of many cells is only one cell thick. The blue growths are rhizoids, with oblique cross-walls. Note that there is variation in the genus. In some Sphagnum species the thallose protonema is ribbon-like and in others it is highly lobed. There may be secondary thallose protonemata which develop at the end of a short filamentous outgrowths from the initial protonema.
In most mosses the protonema is filamentous but in a few genera it is thalloid. The diagram (right) shows an example from the genus Sphagnum. There is a short, filamentous chloronema but it soon develops into a thallose form. The thallose area, though composed of many cells is only one cell thick. The blue growths are rhizoids, with oblique cross-walls. Note that there is variation in the genus. In some Sphagnum species the thallose protonema is ribbon-like and in others it is highly lobed. There may be secondary thallose protonemata which develop at the end of a short filamentous outgrowths from the initial protonema.
 The Hedwigia-type of development is shown by the species Hedwigia ciliata. The spores of this moss have thick walls and the spore divides within the spore wall leading to the existence of two cells within the stretched spore wall. Ensuing growth leads to the rupture of the stretched spore wall and the development of a protonemal component consisting of between 20 and 25 globose cells. This is called a massive protonema and this form of development is shown in the next diagram (right), with some dark grey spore wall fragments visible on the massive protonema. There is no chloronema and the caulonema develops from the massive protonema. The leafy gametophyte buds may develop on either or both of the massive protonema and caulonema.
The Hedwigia-type of development is shown by the species Hedwigia ciliata. The spores of this moss have thick walls and the spore divides within the spore wall leading to the existence of two cells within the stretched spore wall. Ensuing growth leads to the rupture of the stretched spore wall and the development of a protonemal component consisting of between 20 and 25 globose cells. This is called a massive protonema and this form of development is shown in the next diagram (right), with some dark grey spore wall fragments visible on the massive protonema. There is no chloronema and the caulonema develops from the massive protonema. The leafy gametophyte buds may develop on either or both of the massive protonema and caulonema.
The Drummondia-type of development is similar to the Hedwigia-type, except that the massive protonema, of 8-20 cells, occurs within a stretched but unruptured spore wall and there is no caulonema. Since the development takes place within the spore wall it is called endosporous germination. By contrast, the other examples given above show exosporous germination since all protonemal development takes place outside the spore.
Exosporous and endosporous development occur in the liverworts and hornworts as well but the caulonema is found only in the mosses. In the mosses and hornworts exosporous germination is far more common than exosporous but in the liverworts endosporous germination is more common. Liverworts show a variety of protonemal forms (filamentous, ribbon-like, discoid, massive) but liverwort protonemata are generally insignificant in extent, as are those of hornworts![]() .
.
Further gametophyte growth
Growth of bryophyte gametophytes is under control of apical cells. These are cells that divide repeatedly to form new cells. An apical cell has a forward face and up to four rear faces. The rear faces are connected to the rest of the gametophyte but the forward face is free. New cells form only along the rear faces. The newly formed cells are small to start with but then expand so the gametophyte extends in the direction of the apical cell's forward face. Apical cells are fundamental to gametophyte growth. Each stem, leaf, thallus branch and so on has its apical cell or cells. In a leafy bryophyte the leaves grow off stems. At the growing tip of each stem there is one apical cell. There is also an apical cell at the apex of each growing leaf. In thallose bryophytes there may be several apical cells, side by side, in a short row at the thallus margin. The APICAL CELLS CASE STUDY goes into more detail.
Gametophytes produce the male sperm and female eggs. The sperm are produced within antheridia and the eggs are produced in archegonia. Depending on the species in question, both the antheridia and archegonia may be produced on the same gametophyte or on separate gametophytes. In the first case, with both types of sex organs on one gametophyte, the gametophyte is bisexual (or monoicous). In the second there are separate male and female plants, so each gametophyte is unisexual (or dioicous). There are many dioicous species and many monoicous species. Within the monoicous bryophytes the antheridia and archegonia may be on the same or different branches or thallus lobes, again depending on species. People with some botanical or zoological knowledge may have come across the similar-looking terms dioecious and monoecious. You'll find an account of the history and the varied usages of all four terms in the -OICY CASE STUDY. In that link you'll also find descriptions of, and terms for, the various arrangements of sexual organs in the monoicous species. That discussion is put there for those interested in more detail, but it is not necessary for a basic understanding of bryophytes. The most important thing is simply to be aware that there are unisexual and bisexual species.
 Gigaspermum repens |
In dioicous species the male and female gametophytes may be of much the same size or with males smaller, sometimes dramatically so. In this photo of the moss species Gigaspermum repens you can easily see the large-leaved female plants with their spore capsules. The male plants are the much smaller plants that are more easily seen towards the margins of the photograph. This is by no means the most remarkable size disparity, for there are moss species in which the male plants are no more than tiny appendages on the leaves of the female plants.
Some interesting links
The website of the Departamento de Biología de Organismos y Sistemas of the Universidad de Oviedo (in Asturias, Spain) has beautifully drawn representations of the life-cycles of three bryophytes. The captions are in Spanish but should be relatively easily understood by people without a knowledge of Spanish. Many of the technical terms are similar in other languages and there would be few words you'd need to look up in a bilingual dictionary. Here are specific links to the three diagrams:
Phaeoceros laevis, a hornwort.
.JPG){kind=link}
.JPG){kind=link}
.JPG){kind=link}